VARIÉTÉS : UNE SÉLECTION PERMANENTE
L’amélioration génétique de la luzerne vise à produire de nouvelles variétés, productives et de bonne qualité, mieux adaptées aux conditions de culture (climat et itinéraires techniques), aux risques sanitaires (maladies, ravageurs) et aux modes d’utilisation. Elle est basée sur l’exploitation de la variabilité génétique et tient compte de la biologie de l’espèce.

L’autotétraploïdie
Chaque chromosome se trouve présent en quatre exemplaires dans le génome. Chaque individu peut donc porter jusqu’à quatre allèles différents à chaque locus. Cette variabilité allélique constitue une réserve de diversité, mais elle conduit à deux inconvénients : l’identification des combinaisons génétiques favorables et donc le progrès génétique sont ralentis, et des allèles défavorables récessifs sont conservés.
L’allogamie
Les ovules d’une plante sont pollinisés par le pollen d’une autre plante. Le transport du pollen est assuré par des insectes pollinisateurs – surtout des abeilles. En absence de pollinisateurs ou si les fleurs sont peu attractives, la production de graines est faible. Les phases d’autofécondation, souvent utilisées dans les schémas de sélection des lignées ou hybrides pour éliminer les allèles défavorables sont proscrites.
Contrôle des croisements
Dans les parcelles de production il est impossible de contrôler les croisements. Les variétés sont des populations synthétiques issues de plusieurs générations de multiplication de parents bien identifiés. Les parents sont soit des plantes individuelles, soit, le plus souvent des familles de plantes. Plus une variété a de parents, plus sa base génétique est large (plus la variabilité dans la variété est grande). Une diversité fonctionnelle (sur des caractères morphologiques ou physiologiques) pourrait être utile pour améliorer l’adaptation des variétés à des conditions environnementales variées.
En raison de ces particularités, les variétés cultivées sont génétiquement complexes. Toutes les plantes présentes dans un peuplement de luzerne sont génétiquement différentes les unes des autres même si elles sont apparentées. À l’opposé, dans une variété de maïs ou de blé, tous les individus sont semblables, soit tous hybrides (croisement de deux lignées), soit tous de lignées homozygotes que l’on peut reproduire à l’identique par autofécondation.
Néanmoins ces structures génétiques présentent un intérêt majeur : il est bien rare de ne pas trouver des individus portant des allèles favorables dans une variété, lorsqu’un nouveau critère de sélection est travaillé. Ce fut le cas pour la résistance aux maladies : il existait un petit pourcentage de plantes résistantes au sein des variétés sensibles, ce qui a permis de créer de nouveaux pools génétiques améliorés sans utiliser de ressources génétiques « exotiques » qui auraient apporté d’autres caractéristiques défavorables.
La condition préalable à toute sélection est l’existence d’une diversité génétique pour le caractère que l’on cherche à améliorer au sein de l’espèce.
La luzerne cultivée (Medicago sativa) est une espèce fourragère pérenne, originaire du Croissant Fertile (Moyen-Orient). Elle fait partie d’un complexe d’espèces, comprenant à la fois des types diploïdes et des types tétraploïdes. Les variétés cultivées sont toutes tétraploïdes. À ce niveau de ploïdie, les variétés cultivées actuellement sont proches de la sous-espèce sativa, à fleurs violettes, gousses enroulées, port érigé et racine pivotante. Cependant, la sous-espèce sauvage falcata caractérisée par des fleurs jaunes, des gousses en forme de faucille, un port rampant et des racines fasciculées, se croise avec la sous-espèce sativa.
La luzerne cultivée a été introduite en Europe de l’Ouest à l’occasion des grands mouvements de conquêtes romaines puis arabes et redécouverte pour sa production fourragère vers le XIVe siècle. Elle a été introduite plus récemment en Amérique du Nord et en Amérique latine.
La diversité des conditions pédoclimatiques ainsi que la diversité des courants de migration dont sont issues les populations a permis de générer au sein de l’espèce une diversité génétique très grande pour la plupart des caractères d’intérêt agronomique.
Les groupes de diversité génétique dont on dispose en Europe sont d’une part le matériel Nord, (ou «flamand») et d’autre part le matériel Sud (ou «méditerranéen») dont les types «Provence». Les types Nord sont issus de l’introduction à partir des populations sauvages de la sous-espèce falcata de caractères d’adaptation comme la résistance au froid dans la sous-espèce sativa. Une grande diversité existe aussi dans du matériel peu ou pas dormant originaire du pourtour du bassin méditerranéen, de la péninsule arabique et du sous-continent indien. Il se caractérise en particulier par des repousses rapides après les coupes et tolère mieux les coupes fréquentes. Cependant ce matériel est une source de diversité moins utilisée en Europe de l’Ouest en raison de sa faible dormance automnale et donc de sa faible pérennité sous des climats à hivers froids.
Les objectifs de sélection en luzerne sont liés à l’attente des utilisateurs, déshydrateurs ou auto-consommateurs.
La production et la répartition du rendement
La production de biomasse aérienne et sa répartition sur l’année constituent un objectif majeur de sélection. L’évaluation du potentiel de production de matière sèche est conduite en petites parcelles semées à une densité habituelle de culture, dans des dispositifs multi-locaux. Les marges de progrès sur ce caractère sont limitées puisque le produit récolté est directement issu du rendement de la photosynthèse. Cependant, il reste primordial d’adapter le type variétal à la période de végétation de chaque région : privilégier des variétés à faible dormance automnale dans les régions du sud, qui exploiteront les potentiels de production de fin d’automne et de début de printemps ; choisir des variétés assez dormantes dans les régions plus froides, pour ne pas risquer des pertes de plantes à cause du gel. De plus, tous les progrès liés à des tolérances à des maladies se traduisent in fine par des gains de rendement fourrager.
L’adaptation au milieu
La tolérance aux contraintes physiques du milieu passe d’abord par une adaptation de la dormance du matériel génétique au milieu. Cette gestion permet de trouver le meilleur compromis entre la pérennité et le potentiel de production. La dormance automne, qui induit un repos végétatif en période froide, est une protection contre les dégâts de gel. Les variétés dormantes sont probablement les plus sensibles à la durée de la photopériode.
L’adaptation aux conditions de milieu passe aussi par une bonne résistance à la verse, ce qui constitue depuis de nombreuses années un objectif majeur de la sélection française et européenne. La majorité des variétés inscrites au catalogue français présente une bonne tolérance à la verse, même si celle-ci peut être mise en défaut dans des conditions particulièrement humides ou à la faveur de récoltes tardives.
La tolérance aux maladies et parasites
C’est sans conteste le domaine où les progrès génétiques ont été les plus importants au cours des dernières décennies. Les parasites pris en compte sont d’une part les champignons, essentiellement Verticillium albo-atrum et Colletotrichum trifolii dans les zones nord ainsi que Phoma medicaginis en zone sud. Un effort important et fructueux a été conduit pour la résistance au nématode des tiges (Dytilenchus dipsacii) aboutissant à l’inscription de variétés avec de bons niveaux de résistance au cours des dernières années. Des tests existent aussi pour améliorer la tolérance au puceron du pois (Acyrthosiphum pisum), à la fusariose (Fusarium oxysporum) ou à la sclérotiniose (Sclerotinia trifoliorum) mais sont peu utilisés dans les programmes de sélection français. Pour tous ces ravageurs, les tests sont conduits en conditions contrôlées. Des sources de résistance à ces différents parasites ont été identifiées dans les différentes populations disponibles en collection. Certains ravageurs ne sont pas pris en compte en sélection, soit parce que leur impact sur les luzernes est très limité géographiquement, ou que les attaques sont difficiles à reproduire en conditions contrôlées. C’est en particulier le cas du rhizoctone violet.
On notera aussi que certains ravageurs particulièrement virulents sur d’autres zones de culture de la luzerne sont absents ou sans conséquence néfaste en Europe. C’est le cas de Corynebacterium ou de Aphanomyces euteiches.
Teneur en protéines et digestibilité
La qualité du fourrage fait l’objet de travaux de sélection, en vue d’une part d’augmenter la teneur en protéines, et d’autre part d’améliorer la digestibilité du fourrage qui peut être un frein à son incorporation dans des rations pour ruminants à haut niveau de performance. Pour ces deux caractères, la sélection tient compte de la physiologie de la plante, donc de la relation entre croissance et valeur alimentaire. Cela a permis de montrer que pour une production de matière sèche donnée, il existe des différences entre variétés pour la teneur en protéines et pour la digestibilité du fourrage. De plus, une grande variabilité à l’intérieur des populations existe pour ces caractères.
Les caractéristiques de la valeur alimentaire sont mesurées en laboratoire par des analyses chimiques (teneurs en protéines et en fibres, digestibilité enzymatique) et surtout par des prédictions obtenues à partir de spectres collectés dans le proche infrarouge. Des équations de prédiction sont alors préalablement établies, qui permettent de relier l’absorbance à certaines longueurs d’ondes avec des caractéristiques biochimiques ou d’utilisation.
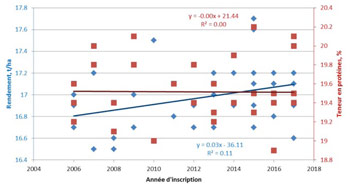 Le développement d’équations de prédiction dans le proche infrarouge permet d’augmenter le nombre d’échantillons analysés dans les schémas de sélection tout en diminuant le coût d’analyses.
Le développement d’équations de prédiction dans le proche infrarouge permet d’augmenter le nombre d’échantillons analysés dans les schémas de sélection tout en diminuant le coût d’analyses.
L’appétibilité de la luzerne a toujours été considérée comme bonne. Cependant, la structure des tiges, et notamment leur grosseur, peut conditionner la consommation en foin par les petits ruminants (ovins, caprins). Dans ce cadre, des variétés à tiges fines ont été sélectionnées, en veillant à maintenir en même temps une bonne résistance à la verse. On a pu montrer qu’il n’existait pas de relation entre la grosseur des tiges et la digestibilité, ni d’ailleurs entre la digestibilité et la résistance à la verse.
Il faut noter que certaines variétés plus récemment inscrites combinent des progrès sur le rendement en biomasse avec un maintien de la qualité. Cela montre que malgré la corrélation négative entre rendement et qualité, les sélectionneurs parviennent à combiner des deux types de caractères.
La production de semences
La production de semences ne fait pas partie intégrante de la valeur agronomique des variétés mais elle contribue à la diffusion du progrès génétique. En effet, elle conditionne la mise en vente des semences à des prix compétitifs. Un critère de sélection efficace, le poids de graines par inflorescence, évalué sur des plantes individuelles, permet d’obtenir rapidement un progrès génétique significatif.diction sont alors préalablement établies, qui permettent de relier l’absorbance à certaines longueurs d’ondes avec des caractéristiques biochimiques ou d’utilisation.
Piétinement et météorisation
D’autres caractères, qui pourraient présenter un intérêt en sélection, sont peu voire ne sont pas pris en compte en Europe.
Parmi ces caractères, on peut tout d’abord citer l’adaptation au pâturage. Cette adaptation recouvre deux aspects que sont : 1. la tolérance des plantes à être consommées directement par les animaux et à supporter le piétinement, sans réduction du potentiel de rendement en biomasse, 2. la gestion du risque lié à la météorisation.
Ce second point passe pour partie par la sélection de génotypes avec une dégradation plus lente des protéines mais surtout par une gestion du pâturage. En revanche, la tolérance à l’arrachement et au piétinement repose sur une modification de la structure des plantes et la recherche de génotypes ayant un port plus prostré. L’aptitude à l’association avec des graminées est matière à de nouvelles études. Dans le contexte de la recherche d’une agriculture durable, les associations luzerne – graminées (dactyle ou fétuque élevée) présentent de nombreux avantages : forte production de biomasse, répartition du rendement plus étalée au cours de l’année, valeur alimentaire équilibrée, réduction de l’utilisation des désherbants chimiques, non utilisation d’engrais azotés de synthèse. Des travaux récents montrent que le classement des variétés diffère sensiblement si on considère le rendement en culture pure ou le rendement en association. Ces résultats sont en faveur de la prise en compte de la valeur en association dans les processus de sélection et d’évaluation des variétés.
La tolérance à la sècheresse et aux stress abiotiques (pH acides, sel, température) font l’objet de travaux de recherche dans différents laboratoires en réponse à des contraintes environnementales très fortes dans certaines régions du globe. Précisément, la réponse à la température (entre 5 et 35°C) de génotypes de type Nord et de type Sud est très semblable pour les processus de croissance et de développement, avec une température optimale entre 25 et 30°C. Néanmoins, les types Sud montrent plus fréquemment des symptômes de stress thermique à 35°C que les types Nord. De façon plus globale, la mesure du rendement, de la qualité et de la pérennité dans des réseaux d’essais donne une information générale sur l’adaptation à une large gamme de contraintes environnementales.

Les variétés mises sur le marché sont issues d’un polycross d’intercroisement entre des plantes ou des familles de plantes sélectionnées pour les critères présentés précédemment. Ce polycross est ensuite suivi de trois ou quatre générations de multiplication de semences avant de parvenir à la semence commerciale.
La spécificité des espèces allogames tient au fait de sélectionner des individus dont les descendants, après différentes générations de multiplication, doivent présenter une bonne valeur agronomique en peuplement dense. Ceci signifie que l’évaluation et la sélection des plantes (ou familles de plantes) parentales effectuées en plantes isolées (pépinière) doivent permettre de prédire le comportement en couvert dense. Pour cela, les descendances de ces plantes sont testées en parcelles denses de petite taille (micro-parcelles), puis les meilleures descendances ou familles feront l’objet d’une évaluation en parcelles plus grandes et dans un dispositif comportant plusieurs lieux. La luzerne étant une espèce pérenne, avec de multiples coupes annuelles, tous ces dispositifs font l’objet de plusieurs récoltes par an et cela sur plusieurs années (en général, l’année d’implantation et les deux années suivantes).
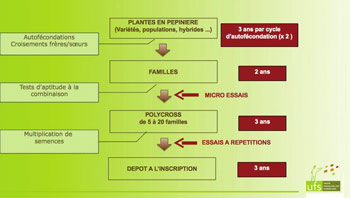 Les conséquences de cette démarche sont en premier lieu le grand nombre d’années pour aboutir à la création de nouvelles variétés. D’autre part, les progrès sont d’autant plus rapides qu’il existe une relation solide entre l’expression du caractère en plantes isolées et l’expression en parcelles denses. C’est le cas pour la résistance à la verse, ou pour la résistance aux maladies évaluée en conditions contrôlées. Quand la relation est plus vague, soit par un effet de la densité sur l’expression du caractère, soit par l’imprécision de la mesure quand elle est réalisée sur des plantes individuelles, les progrès génétiques sont plus lents, comme pour le rendement.
Les conséquences de cette démarche sont en premier lieu le grand nombre d’années pour aboutir à la création de nouvelles variétés. D’autre part, les progrès sont d’autant plus rapides qu’il existe une relation solide entre l’expression du caractère en plantes isolées et l’expression en parcelles denses. C’est le cas pour la résistance à la verse, ou pour la résistance aux maladies évaluée en conditions contrôlées. Quand la relation est plus vague, soit par un effet de la densité sur l’expression du caractère, soit par l’imprécision de la mesure quand elle est réalisée sur des plantes individuelles, les progrès génétiques sont plus lents, comme pour le rendement.
Il faut distinguer deux grandes catégories de technologies susceptibles de faire évoluer la sélection de la luzerne : les techniques de marquage moléculaire et les techniques s’appuyant sur la transgenèse, même si, à ce jour, peu ont eu une application effective sur la luzerne.
Marquage moléculaire :
des travaux aujourd’hui pour demain
Le marquage moléculaire consiste, à l’aide de différentes techniques, à analyser la séquence de l’ADN du noyau ou des organites cellulaires (mitochondries ou chloroplastes). L’abaissement des coûts de séquençage permet désormais de développer de très nombreux marqueurs (plusieurs milliers ou dizaines de milliers) et de génotyper un grand nombre d’individus. Ces outils sont en phase d’évaluation, que ce soit pour les analyses de diversité génétique ou pour aider à l’analyse fine du déterminisme génétique des caractères (identifier les gènes et les allèles favorables).
Ainsi, les marqueurs moléculaires ont permis de mieux décrire la diversité génétique de la luzerne. Ils confirment l’importante variabilité intra-variétale, provenant du mode de reproduction allogame et de la structure synthétique des variétés. En utilisant un grand nombre de marqueurs moléculaires, on distingue néanmoins les différentes variétés, et la structuration rejoint celle observée avec des caractères phénotypiques. Outre ces aspects descriptifs, néanmoins utiles pour gérer un programme de sélection, les marqueurs sont aussi utilisés pour trouver les zones du génome impliquées dans les caractères agronomiques. A terme, l’enjeu est de combiner une sélection phénotypique à une sélection par les marqueurs (sélection assistée par marqueurs). La cartographie génétique des caractères mais surtout la génétique d’association et bientôt la sélection génomique sont les méthodes actuellement étudiées. Il est aussi possible d’étudier la diversité allélique présente pour un gène candidat impliqué dans la variabilité pour un caractère agronomique, de façon à identifier les allèles à effet positif. L’implémentation de ces techniques dans les schémas de sélection dépendra de leur coût et du bénéfice obtenu en terme de progrès génétique par unité de temps. Sur cette espèce autotétraploïde, on peut penser que la création, par sélection assistée par marqueurs, de variétés dont tous les individus portent un allèle favorable sur leurs quatre chromosomes, pourrait permettre un progrès génétique beaucoup plus rapide que par sélection phénotypique.
Ogm : pas en Europe
La transformation par transfert indirect de gènes étrangers permet la création d’organismes génétiquement modifiés (OGM). La luzerne, parce qu’elle est sensible à Agrobacterium tumefaciens peut être transformée génétiquement. De plus, son aptitude à l’embryogenèse somatique permet de régénérer avec une certaine facilité des plantes transformées, même si un petit nombre de génotypes se prêtent avec succès à cette méthodologie. Dans la pratique, peu d’OGM de luzerne ont été effectivement obtenus et testés. La plupart l’ont été à des fins de recherche, en particulier pour l’étude de la fixation symbiotique, et dans des conditions très contrôlées. Des variétés de luzerne « RoundUp Ready » résistantes au glyphosate sont autorisées aux Etats-Unis et au Canada depuis 2013 et 2014. Une autre variété américaine de luzerne, améliorée pour la digestibilité grâce à l’inactivation d’un gène de lignification, a été obtenue en 2014. Pourtant les possibilités sont multiples, tant en ce qui concerne la qualité que la résistance aux maladies ou l’utilisation de la luzerne à des fins de production d’enzymes industrielles.
A côté de la transgenèse « classique », l’édition des génomes à l’aide de l’enzyme CRISPR-CAS9 permet aussi de modifier spécifiquement la séquence d’un gène ou d’introduire un nouveau gène. Cette technologie de précision (« couteau suisse ») est scientifiquement prometteuse, y compris pour la luzerne. Les plantes modifiées par cette méthodologie entrent dans la catégorie des plantes OGM soumises à réglementation, comme en a décidé la Commission Européenne en juillet 2018.
L’acceptation par le marché des OGM est particulièrement faible en ce moment en Europe, et les produits existants n’apportent pas de progrès susceptibles de vaincre ce frein. De plus, il existe en France et en Europe des populations naturelles sauvages compatibles avec la luzerne cultivée ainsi que de nombreuses populations échappées de culture (bords de route). La culture d’OGM entraînerait une dissémination rapide des transgènes dans ces populations spontanés ou sub-spontanées. En conséquence, il n’existe aucun produit OGM qui fasse l’objet d’études au champ en France ni en Europe.