UNE TRÈS BELLE PHYSIOLOGIE
Il est nécessaire de bien connaître et comprendre la physiologie d’une plante pour optimiser sa culture et sa transformation industrielle. Les méthodes de culture doivent en effet tenir compte des particularités biologiques de l’espèce concernée. C’est notamment le cas de la luzerne, dont les applications alimentaires sont variées.
Chez les espèces fourragères, on récolte la biomasse primaire aérienne directement issue de la photosynthèse. Celle-ci a lieu dans les organes riches en chlorophylle, essentiellement les feuilles. La biomasse primaire dépend du rayonnement disponible, de l’aptitude de la culture à intercepter le rayonnement solaire et de son efficacité à utiliser cette énergie solaire dans son métabolisme.
Le rayonnement solaire disponible varie d’un lieu à l’autre, d’une année à l’autre mais on ne peut pas le contrôler. Seule une partie de celui-ci est absorbé par les plantes pour permettre la photosynthèse et cette partie correspond à certaines longueurs d’onde du spectre (essentiellement les bleus et les rouges). L’aptitude de la culture à intercepter le rayonnement incident dépend de son indice foliaire, défini comme la surface totale de feuilles (en m²) par unité de surface de sol.
L’efficience de conversion du rayonnement en produits carbonés est, elle, constante au sein d’une espèce. La luzerne fait partie de la famille des fabacées (ou légumineuses) qui ont une efficience de conversion relativement basse, que l’on peut pour partie expliquer par le coût métabolique que représente la fixation de l’azote par les nodosités des racines. L’aptitude de la luzerne à fixer l’azote atmosphérique représente un atout agronomique majeur pour cette espèce qui peut donc être cultivée sans apport d’azote minéral. Les mécanismes physiologiques de la fixation permettent en outre à la plante d’absorber prioritairement l’azote minéral si celui-ci est disponible, faisant ainsi de la luzerne un excellent piège à nitrates (voir aussi l’article sur les épandages page 56). De ce fait, la luzerne cumule deux avantages : sa culture ne dépend pas de la fabrication d’engrais de synthèse et les fuites d’azote dans l’environnement sont limitées grâce à sa culture.
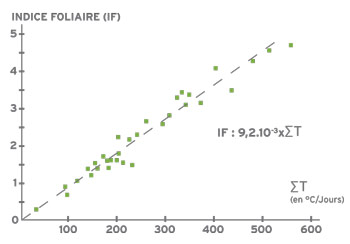 En conditions agronomiques non limitantes (en particulier, nutrition hydrique et minérale), l’indice foliaire évolue de façon linéaire avec les sommes de température reçues par les plantes à partir de la coupe.
En conditions agronomiques non limitantes (en particulier, nutrition hydrique et minérale), l’indice foliaire évolue de façon linéaire avec les sommes de température reçues par les plantes à partir de la coupe.
Ainsi, dans des périodes de températures moyennes faibles, la reconstitution de l’indice foliaire sera lente après une coupe et limitera la production de biomasse. Ceci explique aussi les variations entre les régions pour la rapidité de la reconstitution du couvert.
Il faut un indice foliaire de 3 pour que le couvert intercepte 90% du rayonnement incident.
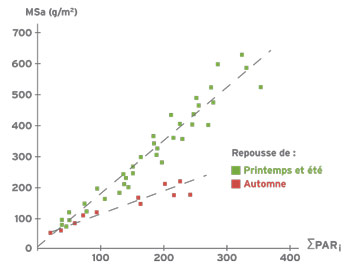 La biomasse aérienne est reliée de façon linéaire à la quantité de rayonnements photosynthétiquement actifs absorbés par le couvert. Cependant, on note une grande différence entre les coupes de printemps et d’été et les coupes d’automne. Pour une même quantité de rayonnements absorbés par un couvert, la biomasse aérienne produite est beaucoup plus faible en automne. Ceci est lié à l’accumulation importante de réserves carbonées dans les racines au cours de l’automne : les assimilats issus de la photosynthèse sont alloués en plus grande proportion aux racines en automne que pendant les autres saisons.
La biomasse aérienne est reliée de façon linéaire à la quantité de rayonnements photosynthétiquement actifs absorbés par le couvert. Cependant, on note une grande différence entre les coupes de printemps et d’été et les coupes d’automne. Pour une même quantité de rayonnements absorbés par un couvert, la biomasse aérienne produite est beaucoup plus faible en automne. Ceci est lié à l’accumulation importante de réserves carbonées dans les racines au cours de l’automne : les assimilats issus de la photosynthèse sont alloués en plus grande proportion aux racines en automne que pendant les autres saisons.
On observe peu de variation entre les variétés disponibles en France pour la vitesse de mise en place du couvert après une coupe. Les seules sources de variation correspondent à différentes classes de dormance.
Les types moins dormants (qui présentent un arrêt de croissance en hiver plus court) ont en général une
reconstitution du couvert plus rapide après la coupe que les types dormants. Cependant, ce matériel végétal est à utiliser avec prudence dans les conditions agricoles
françaises en raison des risques de gel au cours de l’hiver. Les marges de progrès pour la vitesse de repousse sont donc faibles au sein d’une classe de dormance. Néanmoins, le changement climatique avec des hivers moins rigoureux permet d’envisager l’utilisation de variétés un peu moins dormantes.
Le manque d’eau réduit la production de biomasse, en limitant à la fois la restauration de l’indice foliaire et l’efficience de conversion. Il est à noter que la résistance à la sécheresse de la luzerne est toujours très élevée sous nos climats, grâce à un système racinaire très profond, la survie de la luzernière n’étant pas compromise.
Il faut toutefois souligner que l’indice foliaire peut être limité par d’autres facteurs que la température ou la nutrition hydrique : maladies, ravageurs ou tout simplement vieillissement (sénescence) des feuilles.
 Les deux principales composantes de la qualité de la luzerne sont la teneur en protéines et la valeur énergétique. Les protéines sont essentiellement présentes dans les feuilles, il s’agit pour 90% des protéines responsables de la photosynthèse. La valeur énergétique, apportée par les parois cellulaires, est mesurée par la digestibilité ou proportion de biomasse dégradée lors de la digestion, La qualité dépend pour partie de la croissance de la plante.
Les deux principales composantes de la qualité de la luzerne sont la teneur en protéines et la valeur énergétique. Les protéines sont essentiellement présentes dans les feuilles, il s’agit pour 90% des protéines responsables de la photosynthèse. La valeur énergétique, apportée par les parois cellulaires, est mesurée par la digestibilité ou proportion de biomasse dégradée lors de la digestion, La qualité dépend pour partie de la croissance de la plante.
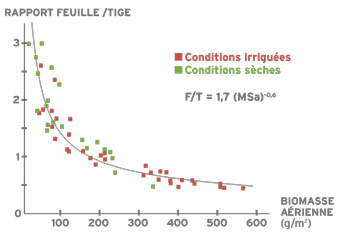
La croissance de la plante se traduit par la production de feuilles et de tiges, support mécanique de ces feuilles. Au cours d’une repousse, le poids de tiges augmente plus vite que le poids de feuilles. Cette évolution de la structure se mesure par le rapport feuille/tige. Ce rapport diminue selon une loi exponentielle en fonction de la biomasse aérienne disponible.
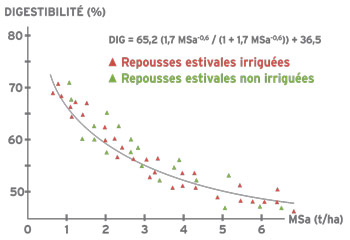 L’évolution du rapport feuilles/tiges a des conséquences importantes sur la qualité du fourrage récolté. Les feuilles, moins riches en parois cellulaires (mesurées par la teneur en fibres), ont une digestibilité plus élevée que les tiges. De plus, la digestibilité des tiges évolue négativement au cours de leur croissance alors que la digestibilité des feuilles reste constante. Sur la base de ces différentes relations, on peut établir la liaison entre la biomasse aérienne produite, le rapport feuilles/tiges et la digestibilité. Pour une variété donnée, le rapport feuilles/tiges est un indicateur de la teneur en fibres et de la digestibilité du fourrage. La réduction du pourcentage de feuilles s’accompagne d’une plus grande teneur en fibres et donc d’une digestibilité plus faible. Il est donc logique que la digestibilité diminue avec l’augmentation de la biomasse aérienne au cours d’un cycle.
L’évolution du rapport feuilles/tiges a des conséquences importantes sur la qualité du fourrage récolté. Les feuilles, moins riches en parois cellulaires (mesurées par la teneur en fibres), ont une digestibilité plus élevée que les tiges. De plus, la digestibilité des tiges évolue négativement au cours de leur croissance alors que la digestibilité des feuilles reste constante. Sur la base de ces différentes relations, on peut établir la liaison entre la biomasse aérienne produite, le rapport feuilles/tiges et la digestibilité. Pour une variété donnée, le rapport feuilles/tiges est un indicateur de la teneur en fibres et de la digestibilité du fourrage. La réduction du pourcentage de feuilles s’accompagne d’une plus grande teneur en fibres et donc d’une digestibilité plus faible. Il est donc logique que la digestibilité diminue avec l’augmentation de la biomasse aérienne au cours d’un cycle.

Les feuilles sont plus riches en protéines que les tiges et la teneur en protéines des feuilles reste assez constante. La teneur en protéines présente une décroissance forte avec l’accumulation de la biomasse, en relation avec la diminution du rapport feuilles/tiges. On peut ainsi passer d’une teneur moyenne en azote de 5% (31% de protéines) avec 1 t/ha de biomasse à une teneur moyenne de 3% (19% de protéines) avec 4 t/ha de biomasse aérienne.
Digestibilité et teneur en protéines diminuent donc avec la croissance des plantes et l’accumulation de biomasse. Cependant, les mécanismes de la décroissance sont différents. Pour la digestibilité, ce sont à la fois l’augmentation de la quantité de tiges et la diminution de la digestibilité des tiges qui interviennent. La teneur en protéines est quant à elle directement liée à la proportion de feuilles dans la biomasse.
L’état hydrique des plantes va jouer un rôle important sur la teneur en azote. Deux processus antagonistes se déroulent simultanément. D’une part, en réduisant la production de biomasse, le stress hydrique contribuerait à produire un fourrage plus riche en azote. Mais, par ailleurs, le stress hydrique réduit la fixation symbiotique, ce qui pénalise fortement la teneur en azote. Ainsi, en cas de stress hydrique, la production de fourrage est moindre et celui-ci est plus pauvre en azote. Les variétés ne sont pas identiques pour leur qualité. Même si la relation entre production de biomasse et qualité de la biomasse reste vraie au cours de la croissance, la sélection pour la digestibilité (par une réduction de la teneur en fibres) et pour la teneur en protéines est active, certaines variétés montrent un progrès pour la production de biomasse avec un maintien de la qualité.
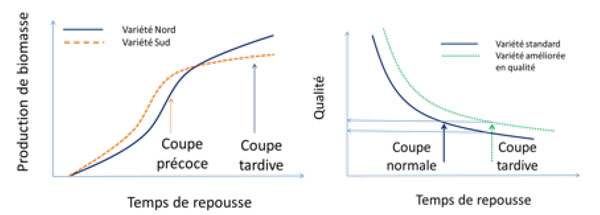
La relation entre la croissance et la qualité est un facteur essentiel de la gestion des calendriers de coupes. Des coupes précoces vont permettre de récolter du fourrage d’excellente qualité, tant pour la teneur en protéines que pour la digestibilité, mais la quantité produite sera faible. De plus, si on adopte un rythme de coupes fréquent, la quantité totale de fourrages produite sur l’année sera plus faible, car après chaque coupe il faut que l’indice foliaire se reconstitue. Si, à l’inverse, on adopte un rythme de coupes lent, on récoltera une quantité de fourrages plus importante mais de qualité plus faible. La gestion des calendriers de coupes permet ainsi d’optimiser la quantité et la qualité de fourrage en fonction des objectifs de l’usine.
 Le système racinaire se caractérise par une racine pivotante puissante capable d’aller puiser l’eau et les éléments nutritifs très profondément dans le sol. Ces racines, et notamment les pivots, représentent une biomasse importante (plusieurs t/ha). Elles montrent des variations de masse au cours d’un cycle de repousse : après la coupe, la plante puise dans ses réserves racinaires carbonées et surtout azotées pour assurer la repousse des tiges.
Le système racinaire se caractérise par une racine pivotante puissante capable d’aller puiser l’eau et les éléments nutritifs très profondément dans le sol. Ces racines, et notamment les pivots, représentent une biomasse importante (plusieurs t/ha). Elles montrent des variations de masse au cours d’un cycle de repousse : après la coupe, la plante puise dans ses réserves racinaires carbonées et surtout azotées pour assurer la repousse des tiges.
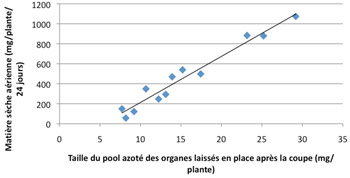
Les réserves carbonées servent principalement à la respiration de maintenance du système racinaire, lorsque l’indice foliaire est insuffisant pour fournir des assimilats issus de la photosynthèse. Par contre, les réserves protéiques accumulées dans le pivot conditionnent très directement la croissance des parties aériennes dans les premiers jours qui suivent une coupe (graphique 7), ce qui a des conséquences sur le rendement en biomasse de la repousse.
Il est impératif que le rythme de coupe respecte le temps nécessaire à la reconstitution des réserves azotées des racines. Si on procède à des coupes trop fréquentes, ou si la nutrition azotée des plantes est déficiente, le système racinaire s’appauvrit en azote remobilisable et la repousse est alors plus faible. Cela conduit à des productions plus faibles puis à la mort des plantes. Ce cycle d’accumulation et de remobilisation vient justifier une ancienne pratique selon laquelle il était nécessaire de laisser fleurir une luzernière au moins une fois dans l’année.